Cytosquelette de Cletini budovi. Cytosquelette. Fonctions du cytosquelette. Cytosquelette de Budov. Crescentine, homologue des protéines des filaments intermédiaires
Les cellules eucaryotes changent de forme, se déplacent, déplacent les organites le long du cytoplasme et séparent les chromosomes lors de la mitose. Tsya zdatnіst est doté d'un maillage trivumirous de fils protéiques (filaments), qui forment l'architecture principale du clitine - le cytosquelette (et il est également désigné comme cytomatrice). Les fibres protéiques imprègnent le cytoplasme des clitines eucaryotes et en l'absence de points d'attache aux protéines de la membrane plasmique de cet organite. Toutes les fibres sont des structures composées de sous-unités - des protéines globulaires spéciales. (Protéines du cytosquelette, comme et d'autres protéines de la clitine, codées dans des gènes et synthétisées sur des ribosomes.)
Les sous-unités du cytosquelette se lient avec des liens faibles (eau, ionim et autres) et ce pouvoir permet aux cellules de façonner l'étendue dynamique de la structure du cytosquelette, qui est facile à modifier. Il est indiqué que dans différentes infusions de clitine, nous changerons le cytosquelette, démontant les principaux composants de son architecture et les formant à nouveau, apparemment jusqu'à la nature du signal supprimé ; dans le même temps, la vie détaillée du cytosquelette change constamment pour la conservation de l'ensemble du plan de l'organisation. Cette forme de système cytosquelettique robotique est appelée principe d'instabilité dynamique.
Selon le diamètre des filaments, ils sont divisés en trois groupes : les microfilaments (5 à 7 nm), les fibres intermédiaires (environ 10 nm) et les microtubules (environ 25 nm). Le type de peau des structures cytosquelettiques est établi dans le clitin par le système énergétique avec ses protéines majeures et mineures. Les systèmes du système sont absolument indépendants et interagissent les uns avec les autres composants de la clitine - la membrane plasmique, le noyau et les autres organoïdes de la clitine. Compte tenu des manifestations évidentes, le cytosquelette n'adhère pas seulement aux formes subcliniques de clitinum, mais aussi à tous les types de clitinite, mais aussi progressivement à une autre partie du clitinum et assure une signalisation sûre au milieu du clitinum vers le larynx un large gamme de complexes protéiques entre les récepteurs et les enzymes.
Microfilaments zustrіchayutsya pratiquement dans tous les types de clitines et est formé à partir de la protéine actine - la plus répandue dans les clitines eucaryotes. (L'actine représente environ 5 % de la protéine cellulaire totale ; dans les viandes squelettiques, environ 20 % de la masse de clitine.) L'actine peut être remplacée par un monomère apparemment (la G-actine est une "actine globulaire" composée de 375 acides aminés résidus) ou des fibres (F -actine - "Actine fibrillaire"). Le filament de cuir F-actine est représentatif d'une structure en spirale d'un petit sprat de micromètres. Les fibres de F-actine peuvent avoir deux charges différentes, elles polymérisent donc avec des tourbillons différents. Un kinet à croissance rapide est appelé un kinet plus, et un kinet à croissance assez rapide est appelé un kinet moins. Plus-kinets croissance du filament d'actine 10 fois plus rapide, moins-kinets inférieurs.
Les microfilaments participent à des processus dynamiques, tels que la stagnation de la muqueuse, les cellules non linguistiques, la phagocytose, l'établissement de la prolifération cytoplasmique dans les cellules rhomomiques et les acrosomes dans le processus d'agrégation des spermatozoïdes avec les ovules. Tous ces processus sont effectués à l'aide de protéines liant l'actine.
Dans le cytoplasme, les clitines ont plus de 50 types différents de protéines de liaison à l'actine, qui interagissent spécifiquement avec la G-actine et la F-actine. Les protéines varient dans différentes fonctions : elles régulent le pool d'actine G (profiline), stabilisent les filaments d'actine F (fragine), cousent les filaments avec d'autres composants du cytosquelette. Les protéines actives de liaison à l'actine, par exemple, la formation de gel (comme le mot - gelée) - grincent les fibres d'actine en croix et, en même temps, transfèrent les cellules du cytoplasme du sol (du latin solutio - rozchin ) dans le gel. Une autre protéine de liaison à l'actine est la spectrine, également appelée fodrine, qui lie les fibres d'actine en faisceaux et les attache à la membrane cytoplasmique et au maillage, qui est induit à partir des fibres proximales. La protéine valine lie les filaments d'actine parallèlement à l'ordonnancement de la même structure et s'intègre dans la douceur de la polymérisation de la G-actine.
Mayzhe tous les types de ruhiv dans le clitiny sont associés à la participation de la myosine, une protéine liant l'actine. Toutes les molécules de myosine ont une tête, un cou et une queue. La tête de myosine est attachée au monomère d'actine I, en raison de la présence d'ATP, s'effondre dans le microfilament plus-moins-moins. Dans les tissus squelettiques, les molécules d'actine et de myosine sont dispersées sur des surfaces de fixation les unes après les autres, et le mouvement des têtes de myosine le long des filaments d'actine entraîne le raccourcissement des membranes. Dans les clitines de langue allemande, lorsqu'ils interagissent avec la protéine myosine, les filaments d'actine peuvent former des faisceaux rapides, les veines sont ainsi établies par invagination (invagination) de la surface de la clitine. De telles invaginations sont établies, par exemple, lorsque rozpodil_ clitin. Zagalom la nature des ruines dans la clitine se trouve sous la forme de la protéine myosine, dont la structure peut être plus de 80 variantes. En combinant des microfilaments d'actine avec différentes variantes de la myosine et d'autres protéines liant l'actine, la clitine forme des structures responsables de l'architecture, du rukhlivistyu et du temps de la muqueuse.
La majorité des microfilaments de clitine soumettent la structure trivimique de la membrane plasmique, appelée cortex d'actine (cortex d'actine). Caractéristiques de la structure de la structure et du renouvellement des microfilaments ; par exemple, dans le cortex d'un leucocyte, les filaments ne durent pas plus de 5 secondes. Le principal type d'interruptions du cortex dans les clitines rukhomi
pseudopodes - prolifération cytoplasmique. Les pseudopodes peuvent prendre la forme d'une plaque plate (lamelopodes), d'un cylindre étroit (phylopode) ou d'un michur buccal. La forme des pseudopodes réside dans le type de protéines de liaison à l'actine, qui interagissent avec les microfilaments et la membrane plasmique.
Les microfilaments d'actine participent au pliage de l'espace ouvert pliable et des structures cytosquelettiques autrement stables. Par exemple, la base des microvillosités des cellules épithéliales de l'intestin et du nirok est formée par les faisceaux de filaments d'actine. Sur la surface supérieure des cellules ciliées, les ravins de l'oreille interne, comme s'ils donnaient lieu à des sons, modifient les spécialisations des excroissances (poils) - stéréocils. Les stéréocils sont disposés en rangées régulières comme les tuyaux d'un instrument de musique à clavier - un orgue. Les stéréocils internes des cheveux vides sont remplis de filaments d'actine et de molécules d'autres protéines. Des mutations de ces gènes, qui codent le nombre de protéines, conduisent à la dégénérescence des cellules ciliées et se manifestent dans l'une des formes de surdité (syndrome d'Usher).
Les microfilaments participent activement à la culture russe. Lorsque les filaments d'actine polymérisent progressivement à l'extrémité du bord rugueux de la clitine, ils se dépolymérisent depuis la face interne. Les processus de polymérisation et de dépolymérisation de la F-actine peuvent être perturbés par des loutres fongiques (toxines). Par exemple, la phaloïdine (toxine du champignon vénéneux) se lie au type négatif de l'actine et inhibe la dépolymérisation, en même temps que la cytochalasine (toxine des moisissures, qui est le pouvoir des cytostatiques) vient au type positif, bloquant le chi polymérisation de l'actine et des cellules ruh. Les discours d'injection de trivalium qui perturbent la polymérisation ou la dépolymérisation des filaments d'actine provoquent la mort de ces cellules.
La polymérisation de l'actine est précisément la régulation du processus, le contrôle des récepteurs superficiels supplémentaires des cellules, des enzymes (protéines kinases) et des ions calcium. La perturbation de ce processus s'accompagne de manifestations cliniques. Par exemple, dans les cellules transformées, il y a un changement dans l'expression des protéines, qui régulent le repliement de l'actine. Des anomalies significatives des filaments d'actine sont observées dans les clitines de certaines bouffées maléfiques. Dans les clitines de sarcome (gonflement des tissus heureux), la présence de filaments d'actine fins et courts a été révélée. La clitine Qi, face à la clitine normale, est plus pourrie et peut avoir une excellente santé avant la métastase.
Filaments intermédiaires sont composés de protéines spécifiques aux types de clitines chantantes (par exemple, les kératines dans les cellules épithéliales, la vimentine dans les cellules des tissus sains, la desmine dans les tissus malins et les tissus). Les filaments intermédiaires donnent le tissu de la clitine, de sorte que la puanteur est micro, fibreuse, résistante à l'étirement du polypeptide et se répand dans tout le cytoplasme de la clitine, satisfaisant le mi du maillage. De plus, des fibres interstitielles sont présentes dans le noyau, satisfaisant le réseau de filaments (lame) sur la surface interne de la membrane nucléaire, qui est étroitement liée aux pores nucléaires.
Les éléments structurels des fibres intermédiaires sont des protéines, qui appartiennent à cinq familles natives et présentent un niveau élevé de spécificité de la clitine. Les représentants typiques de ces protéines sont la cytokératine, la desmine, la vimentine, la gliaprotéine fibrillaire acide et le neurofilament. Toutes les protéines ont une structure de cisaillement de base près de la partie centrale, comme des hélices α. Deux lances peptidiques (dimères) forment une superbobine. Ces dimères se rejoignent de manière antiparallèle, constituant le tétramère. L'agrégation des tétramères selon le principe « tête à tête » donne un protofilament. Vіsіm protofіlamentіv sont épissés à la fois et fibre intermédiaire utavlyuyut d'un diamètre de 10 nm. L'élasticité des filaments intermédiaires est assurée par le fait que les dimères du tétramère dermique sont expansés dans l'ordre de contrôle un par un.
Les cheveux et les ongles des gens, de la laine, des pir'ya, des gols, des pazurs et des hordes de créatures sont formés par le rang de tête de la kératine (cytokératine). Dans une fibre, il y a des millions de fibres entrelacées. Okremi lanceugs la kératine liée à de nombreuses liaisons disulfure, ce qui lui confère une dureté supplémentaire. Plus de 30 kératines différentes ont été observées qui combinent deux types dans les cellules épithéliales humaines. De plus, toutes les isoformes des kératines importantes spécifiques des cheveux et des ongles ont été décrites. Dans les clitines nerveuses, des neurofilaments sont développés pour donner le support mécanique nécessaire aux vieux axones. Filaments d'expansion desminale dans les disques Z des sarcomères malins squelettiques. Dans différents types de clitines, les filaments intermédiaires jouent un rôle important dans la formation des contacts de clitine, appelés desmosomes, qui forment des clitines sucides. Les napivdesmosomes fixent les cellules épithéliales à la membrane basale, de la pourriture puante.
microtubules
Les microtubules présents dans tous les clitines eucaryotes sont de longues structures filiformes, étirées le long de tout le cytoplasme et formant une ligne, améliorant ainsi l'organisation structurelle et la localisation de ces organites.
Les microtubules sont dissous lors de la polymérisation de la protéine tubuline (lat. tubula - tubule), qui est un hétérodimère, dissous par des sous-unités α- і β- tubuline. Au processus de polymérisation α -Tubuline d'un contact dimère β -Tubuline du dimère avancé avec des protofilaments approuvés. Treize rangées tubulaires tardives de protofilaments (fils), qui vont en spirale, établissent un microtubule d'un diamètre de 24 nm et un long sprat de micromètres.
Riz. Schéma de formation des microtubules, qui montre comment les polypeptides de la tubuline, se liant un à un, établissent une paroi cylindrique. A. Coupe transversale B - Coupe courte du microtubule.
Microtubules maillot, pourpointі triolet.
UN un microtubule à un doublet ou un triplet est composé de 13 protofilaments.
tubules Bі C sont constitués d'un plus petit nombre de protofilaments, appelez 10.

La polymérisation des microtubules va directement de la tête à la queue de telle sorte que le microtubule ait la même polarité: les її kintsі sont indiqués comme des kints plus et moins. Les microtubules des clitines sont instables. La puanteur peut rapidement ramasser et ramasser. Dans klittsі minus-kіntsі pov'yazanі z microtubules du centre d'organisation (TSOMT) - structure, noyau roztashovanoї bіlya, aime venger quelques petits corps dans des créatures kіtinakh - tsentriole, utvernih z microtubules, qui étaient en colère. En règle générale, les microtubules sont associés à d'autres protéines (myosine, dynéine, kinésine), reliant ainsi les microtubules à des éléments plus petits du cytosquelette et des organites. Kinezin assure le transport des organites et des vésicules (bulbes) d'une partie du clitinum au point plus du microtubule au point moins, et la dyneine du point moins au point plus.
Vіdomі khіmіchnі spoluki, zdatnі bloquant le repliement des microtubules (colchicine, vinblastine) et stimulant la formation de microtubules stables (taxol). Si les plis des microtubules dans les cellules sont endommagés, alors l'esprit peut être plié (pas trop aigre, température abaissée ou élevée, etc.), si le pliage des microtubules dans les cellules est perturbé, cela peut être causé par un développement normal.
Les microtubules, comme les filaments d'actine, participent à la forme primordiale des cellules. Par ordre de fonction statique, les microtubules participent à divers processus qui se produisent dans tous les clitines eucaryotes : méiose, mitose, rus clitineux et sécrétion. La puanteur est directe "lattes" pour le transport des organites. Avec les protéines associées, les microtubules créent un travail mécanique, par exemple, le transport des mitochondries, le mouvement des bulbes synaptiques, le rux viy (excroissances capillaires de cellules dans l'épithélium de la jambe, des intestins et des oviductes) et la tique de combat du spermatozoïdes. Les pukhirtsі, qui sont établis par l'appareil de Golgi, vont directement aux différentes cellules des cellules le long des microtubules strictement selon les indications. De plus, les microtubules sous forme de fuseau mitotique sont la partie la plus importante de l'appareil, ce qui assure la distribution correcte des chromosomes entre les clitines filles dans la distribution des clitines eucaryotes.
Fonctions des microtubules : 1) assurer la distribution des chromosomes en cas de clivages ; 2) podtrimka forme clitiny; 3) participation au transport des macromolécules et des organites ; 4) assurer la fragilité des flagelles, viy.
Fonctions du cytosquelette
Le cytosquelette a trois fonctions principales.
1. Servir de cadre mécanique du clitiny, ce qui donne au clitiny une forme typique et des liens sécurisés entre la membrane et les organites. Le cadre est une structure dynamique, car il est constamment mis à jour dans le monde des esprits changeants et devient un clergé.
2. Dіє yak "moteur" pour le mouvement du client. Les écureuils Dviguni (courts) se trouvent dans les clitines m'yazovyh et dans d'autres tissus. Les composants du cytosquelette coordonnent directement et coordonnent le mouvement, le rozpodil, le changement de forme de la clitine au cours du processus de croissance, le mouvement des organites et le mouvement du cytoplasme.
3. Servir de "rail" pour le transport des organites et autres grands complexes au milieu de la cellule.
Microfilaments et fibres intermédiaires.
Les microfilaments, stimulés par la F-actine, pénètrent dans les microvillosités, formant des nœuds. Ces microfibres sont combinées avec des blancs supplémentaires, qui sont les plus importants de tous les fimbrins et villins. La calmoduline et l'ATP de type myosine - aza relient les microfibres extrêmes à la membrane plasmique. .
Clitina peut modifier l'ensemble des protéines synthétisées dans le cytosquelette d'une manière en jachère, mais le processus est plus avancé. La conception du cytosquelette peut être facilement modifiée sans synthèse de nouvelles molécules, en raison de la polymérisation et de la dépolymérisation des fils. Dans les cellules, il y a un échange constant de fils et une variété de protéines-monomères dans le cytoplasme. Dans les clitines riches, environ la moitié des molécules d'actine et de tubuline sont situées dans les monomères cytoplasmiques et l'autre moitié pénètre dans le pli du microfilament. Clitina régule la stabilité des fils du cytosquelette, en y attachant des protéines spéciales, qui modifient la stabilité de la polymérisation. Le principe fondamental du fonctionnement du cytosquelette est l'instabilité dynamique. Par exemple, la forme de l'érythrocyte en disque biconcave est soutenue par le cytosquelette membraneux de fibres, titré avec la protéine spectrine. Le spectre de liaison avec la protéine ankérine (ancre - yakir), qui se lie avec la protéine de la membrane cytoplasmique, qui est responsable du transport des anions (Cl - , HCO - 3). Les défauts de la protéine spectrine et de l'ankyrine provoquent la forme non primaire des érythrocytes. Ces érythrocytes s'effondrent déjà rapidement au niveau de la rate. Les maux, yakі vyklikayutsya tels porushennyami, appelés effondrement de la sphérocytose ou effondrement de l'eliptocytose.

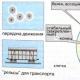
Riz. Cytosquelette des eucaryotes. Les microfilaments d'actine sont transformés en chervonium, microtubules - verts, noyaux de clitine - couleur noire.

Le cytosquelette est composé de trois composants : des microtubules, des microfilaments et des filaments intermédiaires.
microtubules pénétrer tout le cytoplasme de la clitine. La peau d'eux est un cylindre vide d'un diamètre de 20 à 30 nm. La paroi des microtubules est recouverte de 13 fils (protofilaments), torsadés en spirale un sur un. Le fil de peau, avec sa noirceur, est constitué de dimères de protéine de tubuline. La synthèse des tubulines a lieu sur les membranes de l'EPS granulaire et le repliement en spirale se produit au centre de la clitine.
Évidemment, beaucoup de microtubules peuvent être dirigés radialement vers les centrioles. Des bruits de puanteur se répandent dans tout le cytoplasme.
Plus de microtubules peuvent être fixés ("-") et plus ("+") se termine. Vіlny kіnets zabezpechuє podovzhennya qui a tronqué les tubules. viah, ainsi que le centre des chromosomes. En règle générale, les microtubules du cytoplasme gonflent, la puanteur fond au centre de la clitine à une vitesse de 1 µm/min. Microtubules de Ruinuvannya à produire pour modifier la forme des cellules(Tvarinna kіtina nabuvaє forme sphérique sonore). Dans ce cas, la structure du clitin et de l'organite est brisée.
Dans le klitz les microtubules peuvent se dilater :
à la vue des éléments okremih;
en faisceaux, dans lesquels les puants sont liés un à un avec des plaques transversales (excroissances neuronales);
Ø à l'entrepôt de paires ou de doublets (filetage d'essieu de vіy et jgutikіv);
Ø aux triplets d'entrepôt (centrioles et corps basaux).
Dans les deux variantes restantes, les microtubules sont souvent fusionnés un par un.
Fonctions des microtubules :
a) maintenir la forme et la polarité des cellules ;
b) assurer l'ordre du mélange des composants de la cellule ;
c) participation au développement d'autres organites pliables (centrioles, veines fines.);
d) participation au transport intracellulaire ;
e) prévention du déplacement chromosomique en cas de clitinite mitotique à rozpodila;
e) la sécurité du ruhu viy.
Microfilaments. Les microfilaments sont appelés fils fins de couleur blanche d'un diamètre5 - 7 nm, qui sont pratiques dans tous les types de cellules. La puanteur peut se propager dans le cytoplasme en grappes, en boules en forme de maille ou une à une.
La principale protéine des microfilaments est l'actine, souvent jusqu'à 5% de la quantité totale de protéines. Krim nouveau, à l'entrepôt de microfilaments peut inclure la myosine, la tropomyosine, ainsi que des dizaines de protéines de liaison actives. La molécule d'actine ressemble à deux fils torsadés en spirale. Le maillage cortical est froissé sans milieu sous la membrane plasmique, dans une sorte de microfilament s'entrelaçant entre lui-même et un à un derrière l'aide de protéines spéciales, par exemple la filamina. La ligne corticale est formée par la douceur du changement de forme de clitine, changeant pas à pas avec la participation d'enzymes décomposant l'actine. Tim lui-même ne changera pas la déformation aiguë et rapide du clitin avec des injections mécaniques. Les microfilaments de la membrane corticale sont attachés aux protéines intégrales et transmembranaires de la membrane plasmique, ainsi qu'aux couches dites adhésives (contacts focaux), car ils recouvrent le clitina avec les composants de la parole intercellulaire ou avec d'autres clitina tinami. Les microfilaments sont plus résistants aux infusions physiques et chimiques, les microtubules inférieurs.
Fonctions principales des microfilaments :
1) assurer la raideur chantante et l'élasticité du clitin pour le maillage cortical des microfilaments ;
2) modifier la consistance du cytosol, y compris le temps de transition du sol au gel ;
3) participation à l'endocytose et à l'exocytose ;
4) assurer la fragilité des clitines non linguales (par exemple, les neutrophiles et les macrophages), qui est basée sur le changement de forme de la surface de la clitine après la polymérisation régulée de l'actine ;
5) le sort de la clitine et des fibres m'yazovykh à courte durée de vie;
6) stabilisation des plis locaux de la membrane plasmique, qui sont protégés par des faisceaux de filaments d'actine réticulés (microvillosités, stéréocils) ;
7) participation à la formation de spores intercellulaires (re-mastication des démosomes et in).
Les filaments intermédiaires sont tissés avec des fils de corde blancs.zavtovshki près de 10 nm. Un tel pokaznik a zoomé sur l'introduction de la masse intermédiaire entre les microtubules et les microfilaments. Promіzhnі filamenti utavlyayut trivіrіnі merezhі dans les cellules de différents tissus d'un organisme vivant. La puanteur suinte du noyau et peut se trouver dans différents compartiments du cytoplasme, établir des septa intercellulaires (desmosomes et napivedesmosomes), se développer au milieu des cellules nerveuses.
Fonctions principales des filaments intermédiaires :
1) structurel ;
2) soutien ;
3) la fonction de subdivision des organites dans les cellules chantantes de la clitine.
Le cytosquelette est une collection de structures protéiques filiformes - microtubules et microfilaments, qui forment le système de support-ruffle de la cellule. Le cytosquelette ne peut être que des cellules eucaryotes, dans les cellules des procaryotes (bactéries) il n'y en a pas, ce qui est une distinction importante entre ces deux types de cellules. Le cytosquelette du clitin sing est formé pour l'apparition de la paroi dure du clitin. Vient organiser tous les organoïdes dans le cytoplasme (le soi-disant dépassement protoplasmique), qui sous-tendent la ruée amiboïde. Le cytosquelette est facile à changer, sans qu'il soit nécessaire de modifier la forme de la clitine. La construction de clitine modifie la forme du déplacement cumulatif des couches de clitine aux premiers stades développement germinal. Quand rozpodіlі kitini ( mitose) le cytosquelette est "désassemblé" (dissocié), et dans les clitines filles, l'auto-pliage est renouvelé.
Le cytosquelette a trois fonctions principales.
1. Servir de cadre mécanique du clitiny, ce qui donne au clitiny une forme typique et des liens sécurisés entre la membrane et les organites. Le cadre est une structure dynamique, car il est constamment mis à jour dans le monde des esprits changeants et devient un clergé.
2. Dіє yak "moteur" pour le mouvement du client. Les écureuils Dviguni (courts) se trouvent dans les clitines m'yazovyh et dans d'autres tissus. Les composants du cytosquelette coordonnent directement et coordonnent le mouvement, le rozpodil, le changement de forme de la clitine au cours du processus de croissance, le mouvement des organites et le mouvement du cytoplasme.
3. Servir de "rail" pour le transport des organites et autres grands complexes au milieu de la cellule.
24. Le rôle de la méthode d'immunocytochimie dans la perforation du cytosquelette. Particularités de l'organisation du cytosquelette dans les cellules malignes.
L'analyse immunocytochimique est une méthode qui permet de réaliser des analyses immunologiques du matériel cytologique dans l'esprit de la préservation de la morphologie de la clitine. L'ICC est l'un des types anonymes de la méthode immunochimique : dosage immunoenzymatique, immunofluorescence, radioimmunodosage. La base de la méthode ICC est la réaction immunologique à un antigène et à un anticorps.
Le cytoplasme des clitines eucaryotes est imprégné d'un réseau semblable à un trivum de filaments protéiques (filaments), appelé cytosquelette. Selon le diamètre des filaments, ils sont divisés en trois groupes : les microfilaments (6-8 nm), les fibres intermédiaires (environ 10 nm) et les microtubules (environ 25 nm). Toutes les fibres sont des polymères, qui sont formés à partir de sous-unités de protéines globulaires spéciales.
Les microfilaments (filaments d'actine) sont constitués d'actine, une protéine qui est la plus large dans les cellules eucaryotes. L'actine peut être utilisée sous forme de monomère (G-actine, « actine globulaire ») ou de polymère (F-actine, « actine fibrillaire »). La G-actine est une protéine globulaire asymétrique (42 kDa), constituée de deux domaines. Dans le monde du mouvement de la force ionique, la G-actine s'agrège de manière réversible, tordant le polymère linéaire en hélice, la F-actine. La molécule de G-actine porte la molécule d'ATP (ATP), qui est liée à la molécule d'ATP (ATP), lorsqu'elle passe pendant une heure à la F-actine, elle s'hydrolyse en ADP (ADP), tobto. La F-actine montre le pouvoir de l'ATP-ase.
B. Protéines des fibres intermédiaires
Les éléments structurels des fibres intermédiaires sont des protéines, qui appartiennent à cinq familles natives et présentent un niveau élevé de spécificité de la clitine. Les représentants typiques de ces protéines sont la cytokératine, la desmine, la vimentine, la gliaprotéine fibrillaire acide [CFGP (GFAP)] et le neurofilament. Toutes les protéines ont une structure de cisaillement de base près de la partie centrale, comme une hélice α superenroulée. De tels dimères s'associent de manière antiparallèle pour former un tétramère. L'agrégation des tétramères selon le principe « tête à tête » donne un protofilament. Vіsіm protofіlamentіv utvoryuyut fibre industrielle.
À la surface des microfilaments et des microtubules, il est peu probable que les monomères des fibres intermédiaires soient rétrécis près du cytoplasme. La polymérisation conduit à la solubilité de molécules polymères non polaires stables.
V. Tubuline
Les microtubules sont formés à partir de la tubuline, une protéine globulaire, qui est un dimère de sous-unités α et β. Les monomères de tubuline se lient au GTP (GTP), qui hydrolyse ce GDP (GTP). Deux types de protéines sont associées aux microtubules : les protéines structurales, les lk-translocateurs.
La discussion même sur le cytosquelette a été proposée pour la première fois par Koltsov, un éminent cytologiste russe du début du XXe siècle, né dans les années 1920. Les éléments du cytosquelette sont striés dans toutes les clitines eucaryotes, et l'axe des analogues de ces structures est є i chez les procaryotes. Le niveau de diversité des éléments du cytosquelette dans les différents clitines. Par exemple, les cellules de l'épiderme sont particulièrement riches sur les filaments proximaux. Les fibres de viande ont plus de microfilaments d'actine et les microtubules sont plus striés dans la croissance des cellules nerveuses, des cellules pigmentées. Les pouvoirs primordiaux des éléments du cytosquelette sont ceux qui sont des polymères protéiques fibrillaires qui ne se chamaillent pas, s'accumulant jusqu'à une augmentation de la surface et se ruinant. Une telle instabilité des éléments du cytosquelette conduit à la friabilité des cellules. Par exemple, changez leurs formes. Les composants actifs du cytosquelette avec la participation de protéines additives spéciales peuvent stabiliser et établir des ensembles fibrillaires repliables, jouant le rôle de la charpente. Lorsqu'ils interagissent avec d'autres protéines spéciales, qui peuvent être considérées comme des protéines motrices ou des translocateurs, les composants du cytosquelette peuvent participer à divers troubles cellulaires.
Le cytosquelette est constitué de trois sous-systèmes. La puanteur est de se battre pour l'entrepôt, l'ultrastructure, pour les autorités fonctionnelles. Le système de microfilaments (actine-myosine), le système de microtubules (tubuline-dynéine) et le système de filaments intermédiaires (filament de 10 nm).
Microfilaments touffes dans le cytoplasme des cellules en ruine des créatures, donc la boule corticale (sous la membrane plasmique) elle-même, et dans les cellules en croissance et les champignons se développent dans des boules de cytoplasme, qui s'effondrent. La principale protéine de microfilament est la protéine d'actine. Tsé complexe kіlkoh bilkіv. La protéine cutanée de ce complexe est codée par son propre gène. Deux types d'actine sont observés - la forme monomérique (forme globulaire) de la G-actine., qui venge la molécule d'ATP. Lors de la polymérisation de la G-actine, une fine fibrille se forme, d'une taille d'environ 8 microns. Cette structure est appelée F-actine. Les microfilaments d'actine sont polaires pour leur dominance. Ces structures dynamiques, yakі peuvent être sélectionnées et analysées de manière en jachère dans l'actine globulaire et fibrillaire spіvvіdnennia.
Le système fibrillaire instable des clitines est stabilisé par un grand nombre de protéines supplémentaires, qui interagissent avec la F-actine. ainsi, par exemple, la protéine tropomyosine assure le jeu de plusieurs filaments d'actine, leur conférant une dureté. Les protéines filamine et alpha-actinine établissent des liens croisés entre les fils d'actine F, ce qui conduit à la formation d'un maillage trivial pliable. Tsya merezh nadaє gelepodіbnogo Je deviendrai un cytosol. D'autres protéines supplémentaires peuvent lier des faisceaux de filaments. Par exemple, la protéine fimbrina. De plus, des protéines sont utilisées, car elles interagissent avec les microfilaments et empêchent la pourriture. L'interaction de la F-actine avec d'autres protéines supplémentaires régule l'état d'agrégation des microfilaments, les protégeant des peluches ou, navpaki, même de la pourriture. І sécuriser l'interopérabilité avec les autres composants.
J'ai un rôle particulier dans l'interaction avec la myosine, la protéine grise d'actine. Vin n'est pas apporté aux protéines supplémentaires. Vin est un autre composant principal du système d'actine.
La myosine est une famille de protéines similaires. Dans toutes ces protéines, on voit que la structure a une tête et une partie motrice, ce qui indique l'activité du complexe pour l'ATPase. Un autre composant des protéines de myosine est le col de l'utérus, lié au décalcome par des protéines régulatrices. Le troisième composant est la partie de la queue, qui est spécifique au type de peau de la myosine et signifie la fonction de cette protéine.
L'ensemble du complexe de myosines est divisé en trois types : la myosine I, la myosine II et la myosine V.
La myosine I. est une molécule monomère.
Myosini II et V sont des dimères. La partie de la section de queue constitue la structure dite superspirale alpha. 2 molécules de myosine II peuvent interagir entre elles et former une fibrille.
Les myosines I et V participent à l'interaction du cytoplasme de la membrane, par exemple au transport des vésicules. Le mécanisme d'interaction de ces protéines, principales protéines du système des microfilaments, part de l'interaction de la tête de myosine avec le filament d'actine, qui conduit à la disparition des molécules de myosine et à un déplacement à distance.
Au cours du cycle cutané, la tête de myosine se déplace directement de l'extrémité positive du filament d'actine vers l'hydrolyse d'une molécule d'ATP de 5 à 25 nm. Cela est dû au forgeage direct du filament d'actine avec les molécules de myosine. Modèle Tsya nom distal modèle Huxley. Théorie des molécules forgées.
Fibres transversales smugasti m'yazovі є zbіshenoyu modèle mikrofilamentu. Myofibrilles avec un fil de 1-2 microns avec des coupes sombres et claires, qui sont dessinées. Seul, budovi myofibrille є sarcomir ou un dilyanka entre deux disques Z ou blancs. Les fonctions du disque Z sont similaires aux liens entre les structures vasculaires. Les protéines Z elles-mêmes sont des structures courtes.
La propagation des sarcomères à l'état détendu varie de 1,8 à 2,8 microns. Uzdovzh sarcomère roztashovuyutsya trois morceaux de protofibrille. Mince, attaché avec un disque Z, comme des fils d'actine. Je tovstі threads, représenté par des molécules de myosine. Roztashovuyutsya tovstі enfile le yak aux interstitiels entre les fils d'actine.
Les têtes des molécules de myosine interagissent avec les filaments d'actine et forment des complexes actine-myosine à la suite de l'interaction de deux protéines indépendantes. L'activité de ces complexes dans les replis riches est supérieure à celle de l'activité ATPase d'une protéine, la myosine.
La brièveté de la myofibrille est affectée par le changement entre les disques Z. Tobto. La longueur du sarcomère raccourcit d'environ 20 pouces. Le mécanisme de raccourcissement du raccourcissement du raccourcissement coopératif de tous les sarcomères après la longueur des myofibrilles. La base de la brièveté est le mouvement d'un fil fin et épais, avec lequel les fils de myosine pénètrent dans l'espace entre les fils d'actine, en s'approchant du disque Z.
Comment fonctionne le système de microfibrilles dans l'entrepôt du cytosquelette :
1) Tassement de l'appareil à déplacement rapide du clitin, qui assure la fragilité.
2) Moulage des structures squelettiques, s'accumulant jusqu'aux mains mouillées pendant le processus de polymérisation et de dépolymérisation de l'actine (G-actine et F-actine).
3) Mouvement mécanomécanique dans les processus d'endo- et d'exocytose et de cytotomie (rozpodil tila clitinis).
Une autre partie musculo-squelettique du cytosquelette - système de tubuline ou système de microtubules. Le système de microtubules peut être richement cohérent avec le système actine-myosine précédemment examiné. Semblable à elle, en premier lieu, s'accumulant jusqu'à la polymérisation et la dépolymérisation. D'une autre manière, la polarité des fils blancs est également possible. Troisièmement, il existe un grand nombre de protéines supplémentaires.
La principale protéine du système est la tubuline. La tubuline est un hétérodimère. Il se compose de deux parties - la tubuline alpha et bêta. Sous-unité Qi lors de l'association utvoryut tubuline bien protéine.
Au cours du processus de polymérisation, les molécules de tubuline sont combinées de telle sorte que la sous-unité bêta interagisse avec la sous-unité alpha et que la sous-unité alpha interagisse avec la sous-unité bêta.
Ces molécules sont vishikovuyutsya une par une dans un long fil de protofilament.
Parallèlement, lorsque le protofilament est ajusté à une longueur, lors de la polymérisation, on observe une augmentation de la largeur. Dans l'ordre des chèques. Franges jusqu'à un maximum de 13 protofilaments. Les protofilaments tardifs sont tordus dans un tube vide, dans une telle tubuline monomère de peau se caractérise par une taille linéaire de 5 nm. Le diamètre réel du cylindre, qui s'est stabilisé, est d'environ 25 nm. L'axe de ces microtubules, apparu à la suite de la polymérisation de quatre molécules de tubuline dans le cytoplasme, est appelé microtubule unique. Ces structures dynamiques. Instabilité dynamique- Trouver les caractéristiques du tube. La puanteur reprend rapidement et reprend rapidement. Ce processus consiste à établir le spiving de molécules de microtubules simples et organisés dans les cellules.
À une concentration suffisante de tubuline protéique, la polymérisation se produit spontanément et le degré de polymérisation est plus susceptible de se produire dans l'une des terminaisons des microtubules, appelée terminaison positive. Avec une concentration insuffisante de tubuline, les microtubules sont séparés des deux sens. La séparation des microtubules va, d'une part, abaisser la température, et d'autre part, ce processus dépend de la présence d'ions calcium.
Ils y voient un saupoudrage de types de discours, de plantes alcooliques, qui indiquent le caractère aléatoire de la sélection des molécules de tubuline. La colchicine alcaloïde la plus répandue. Tsya speech vzaimodiє z okremi molécules de tubuline et zabigaє polymérisation. L'heure médiane de la vie est environ cinq fois plus ancienne. Un tel camp est pritamanny interfazi. Okremі microtubules sur kіntsi, croissance shcho, podzhuyutsya zі swidkіst 4-7 microns / plume, puis dosit shvidko raccourcir. 14-17 µm/m. Dans les cellules qui se divisent, les microtubules s'assemblent en une structure particulière. Elles sont organisées en un fuseau sous achromatique, qui assure le processus de diffusion du matériel génétique entre les cellules filles. L'heure de vie des microtubules dans l'entrepôt d'un fuseau achromatique n'est que de 15 à 20 secondes. Il est important que l'instabilité des microtubules soit due à l'occlusion de l'hydrolyse du GTP. Cependant, 20% des microtubules sont occupés avec un étirement apparemment stable de 20 ans dans les cellules différenciées. En raison de la stabilité due à la modification de la tubuline.
Les microtubules eux-mêmes ne sont pas de courte durée, ce sont les composants visqueux externes des organites cellulaires qui s'effondrent, tels que le flagelle, le fuseau achromatique ci-dessous, comme les microtubules du cytoplasme, comme les viscosités externes pour le transport cellulaire interne, les processus exocytes oz, endocytose et transport de toutes les espèces.
La micronémie unique cytoplasmique, localisée dans l'hyaloplasme, remplit deux fonctions - squelettique (squelettique) et squelettique rugueuse - dans la stabilisation de la forme clitine. Avec une conception pièce par pièce, la clitina perd sa forme et devient un sac. Construisant des organisations clitoridiennes internes, les microtubules sont les fonctionnaires des structures cellulaires internes orientées vers le mouvement.
Le rôle moteur des microtubules joue un rôle dans le fait que les puants créent un système vectoriel ordonné de mouvement. Les terminaisons positives des microtubules sont redressées du centre de la cellule vers la périphérie. Et la présence d'extrémités polaires à la fois positives et négatives des microtubules avec des dynéines crée la possibilité de transférer des composants dans la clitine de la périphérie vers le centre.
Les microtubules se développent à partir du centre d'organisation des microtubules (MCMT).
Dans ces centres, les microtubules commencent leur croissance dans des plantes spéciales et se développent de manière polaire. La direction positive du microtubule se développe. Comme COMT dans les clitines des créatures, le rang principal prend le sort de la matrice des centres clitines ou centrosomes. Avec leurs extrémités négatives, les microtubules sont ancrés au COMT et ancrés en eux. Sous l'influence de cim, il est nécessaire de comprendre l'interaction avec des protéines spéciales, qui doivent être entourées d'un ensemble de microtubules. Dans les clitines de plus grande croissance, la polymérisation des microtubules s'étend le long de la périphérie du noyau de la clitine, auquel cas les tubules divergent radialement.
Dans la plupart des cas, dans les cellules interphases des créatures, l'organisme nouvellement créé et la croissance des microtubules ressemblent à une lumière spéciale.
1) Les microtubules sont formés en structures organisées entrant dans l'entrepôt de viy, centrioles et flagelles, rux viy et flagelle de chienne.
2) Les microtubules sont organisés en fils d'un fuseau achromatique au fond lorsque les cellules sont séparées.
3) Il stimule le transport des cellules moyennes, la membrane mobile, les protéines de sécrétion et de transport et les organoïdes.
4) Є cytosquelette du clitin, sans besoin de maturation.
Conférence : centre clitine (centrosome)
Le centre centrosome ou clitine a été découvert en 1875 par Fleming. En 1876 - Beneden. Roztashovuyutsya au centre géométrique du clitin. La puanteur est caractéristique des créatures. Il n'y a rien de tel dans les grands roslins, dans les champignons inférieurs et les plus simples. Au centre clitiny, il y a des centrioles dribni schіlnі tiltsya, son à la paire. Une paire de centrioles est un diplosome. Dans cette paire de centrioles, les orientations sont perpendiculaires une à une. Le diplosome est aiguisé avec un cytoplasme léger, avec des fibrilles radialement minces - la centrosphère.
La base du budіvlі tsentrіolі est repliée sur le pieu neuf microtubules de triplets. Il y a neuf triplets d'un cylindre vide d'une largeur d'environ 0,15 micron et d'un fond de 0,3 à 0,5 micron. Le premier microtubule du triplet est appelé a-microtubule. C'est juste un microtubule. L'autre est le troisième microtubule, je ne m'en souviens pas. La puanteur se venge de 11 protofilaments et jouxte vprituly un à un En d'autres termes, un dilyanka qui zadnuє microtubules є zagalnoy.
Le triplet de coutures en cuir est à environ 40 degrés sous le haut du cylindre jusqu'au rayon du cylindre. Les microtubules sont repliés à partir d'une tubuline. La crème de tubuline à l'entrepôt de centriol comprend des structures supplémentaires représentées par la dynéine protéique.
Son, dans les cellules d'interphase près de l'entrepôt de diplosomes, on voit le centre mère et la fille. La fille grandit perpendiculairement à l'axe postérieur du centriole maternel. A la partie centrale du centrole, la soi-disant manche est représentée par la nexine blanche. Nexin a la forme d'un voile, appelé aiguilles à tricoter, neuf aiguilles à tricoter dans un triplet de peau droite. Le volume, qui se trouve au milieu du moyeu central à rayons, peut être pris de 3/4 à 1/5. Une série de diplosomes sous la forme du centriole maternel se dresse en apparence d'une matière amorphe de virosité, comme on les appelle appendices ou satellites du centriole maternel. Les appendices de la fille n'en ont pas.
Le système de microtubules de centrioles est décrit par la formule 9+0. Près des centrioles de la matrice à fibres fines se trouve un embrayage dans lequel les microtubules sont enchevêtrés. Les mufti ont des satellites (= satellites recentriolaires). La puanteur est formée de structures fibrillaires d'un pied en tricot. Le fond porte une tête. Contact avec les autres. Les satellites sont des centrifugeuses sur lesquelles sont repliés des microtubules.
Une telle morphologie de diplosomie n'est pas donnée. Tout est encore une structure plastique. Budov, que l'activité du centrosome change radicalement en jachère pendant la période du cycle de la clitine.
cycle de la clitine l'heure est appelée de l'épi de la crémation de la cellule au її du podіlu humide.
Période : rozpodil (noyau rozpodil et cytoplasme rozpodil), devenu environ 1/7 partie du cycle de la clitine. Et reshta - la période de préparation vers le bas (interphase).
Le stade cutané du cycle de la clitine est caractérisé par ses propres particularités de métabolisme et de morphologie.
Sous l'heure, les cellules possèdent 2 centrosomes. La clitine peut avoir 4 centrioles, la puanteur se répand aux pôles de la clitine comme 2 diplosomes. Le centriole maternel à tous les stades de la mitose est aiguisé avec une large zone, d'environ 0,3 µm de large, représentée par de fines fibrilles. Cette zone est appelée halo fibrillaire centriolaire. Les microtubules pénètrent radialement à travers ce halo. Il est important que le centrole fille n'ait pas de halo ou de microtubule. I un tel diplosome viconise la fonction de former le fuseau de l'appareil mitotique. Broche achromatique.
La zone des diplosomes, la centrosphère des diplosomes, est appelée matrice péricentriolaire, centre d'organisation ou de polymérisation des microtubules (COMT). La première forme d'activité des centrioles.
Centrioles - centres de polymérisation des microtubules. Jusqu'à la fin de la télophase, si le cytoplasme est pratiquement détruit par la clitine, les chromosomes commencent à se décondenser et de nouveaux noyaux filles s'établissent. Le fuseau achromatique s'effondre et les tubes du fuseau se dépolymérisent. Les centrios clitiniques changent également de structure, les centrioles maternel et fille eux-mêmes sont mutuellement perpendiculaires. Variable jusqu'à 2 microns. Pendant la période G1, des satellites se forment sur l'épi, à travers lesquels les microtubules pénètrent radialement. Les centrioles deviennent un milieu pour la formation de microtubules cytoplasmiques. Dans le monde de la croissance, les microtubules sont connectés à la région des centrioles et les microtubules se retrouvent librement dans le cytoplasme pendant une heure. Dans la clitine, il y a un changement de convoyeur et une reproduction des microtubules cytoplasmiques. Dès que les clôtures des cellules passeront en phase offensive, ce sera une phase calme (période G0).
La transition de la clitine au stade de son attachement fonctionne au fonctionnement du centre de la clitine en tant que structure, qui forme soit une membrane plasmique viriste remplie d'un axonème. Axonème - filetage axial.
L'axonème est plié en neuf doublets de microtubules, qui se développent à partir des centrioles, et s'étendent également radialement le long du pieu, et dans le doublet cutané, on peut voir un microtubule irrégulier. La crème est un doublet de microtubules, car viz se caractérise par la présence de deux microtubules centraux simples, en forme de nexine protéique additive sous la forme d'un cylindre axial ou central. (9+2). Centrioli a battu la fonction du corps basal.
À la période actuelle S, le centre clitine gagne une forme d'activité de plus et le nombre de centrioles lui-même augmente. La multiplication des centrioles n'est pas associée à leur rozpodil, mais est formée par l'établissement du rudiment ou procentriole, tel qu'il se forme sur la paroi du centriole évident perpendiculaire au centriole de la peau. Neuf microtubules simples sont posés sur le dos, puis la puanteur se transforme en neuf doublets puis en neuf triplés. Cette extension est appelée duplication. En raison de cette croissance des structures à l'arrière de la tête, un petit centrôle de fille est établi, à mesure que nous grandissons jusqu'à la taille de celle de la mère. Dans la période S, en même temps que la duplication, le centriole maternel continue à établir des microtubules cytoplasmiques.
À la suite du processus de duplication du centriole de la peau, le nouveau centriole se développe. La duplication des centrioles est un mécanisme déclencheur ou un signal de réplication de la molécule d'ADN. Après l'achèvement de la période S, le klitz a déjà changé deux diplosomes.
Vient ensuite le début du cycle de la clitine. La période post-synthétique, qui précède directement le rozpodіlu. Les mêmes connaissent les satellites au diplôme de la mère. Les centrioles maternels offensifs sont recouverts d'un halo fibrillaire et commencent à former maintenant des microtubules mitotiques.
Pourpre, dans le cytoplasme, il y a une rupture des microtubules et de la clitine sous la bonne forme. Klitini, comme la construction d'une reproduction triviale, la puanteur se répète de cycle en cycle. Si la clitine reste à la station G 0 période, alors le centrole participe, d'abord, au processus de polymérisation des microtubules cytoplasmiques, et d'une autre manière, à l'adoption du dispositif pour la circulation des tubules.
Oui subdivisés en deux groupes : les kinétocils, qui sont typiques des cellules épithéliales spéciales, ou des cellules flottantes et des veines primaires.
Via est un viriste cylindrique mince dans le cytoplasme avec un diamètre constant de 300 nm. Viriste de la base au sommet du revêtement avec une membrane plasmique. Au milieu de la croissance, la structure de l'axonème s'est développée, qui est principalement composée de tubuline et de dynéine.
La partie proximale inférieure du vії zanurenny dans le cytoplasme et s'appelle corps basal. Le diamètre axonémique du corps basal est le même. L'axonème a neuf doublets dans son entrepôt, qui constituent la paroi extérieure du cylindre axonémique. Les doublets de microtubules sont légèrement tournés sous l'apex d'environ 10 degrés par rapport au rayon axonémique. Dans les doublets, les microtubules sont divisés également, soit le microtubule A, qui se compose de 13 protofilaments et les microtubules B, pas exactement, il peut y avoir 11 protofilaments. Les microtubules A exercent leur propre force, yakі se dirigeant vers les microtubules B à partir du doublet de suture. Mouler les poignées de la protéine additive à la dynéine. Définition de représentations par de grands complexes protéiques, qui sont composés de 9 à 12 lances polypeptidiques, qui peuvent être remplacées par 2 à 3 têtes globulaires, reliées à la fois par des ligaments linéaires plus elliptiques. Le dyneinu de la tête de la peau peut avoir un centre actif d'interaction avec la molécule d'ATP. Dans le microtubule A au centre du cylindre central, des protéines auxiliaires radiales pénètrent, comme si elles formaient des rayons, qui pénètrent dans le cylindre central.
Le corps basal est le même que le budov, comme le centrole. Les anses, la douille et les rayons, sont cousus dans la partie inférieure du corps basal. Sur l'extension du corps basal, qui jouxte la membrane plasmique, il y a neuf appendices qui traversent le triplet cutané jusqu'à la membrane plasmique et se connectent au tissu de clitine. Par conséquent, le corps basal et la vіya sont structurellement liés un à un et deviennent un tout. Les microtubules A et B dans les triplets du corps basal se poursuivent en microtubules A et B dans les doublets d'axonèmes. Et l'axe de la partie interne de la vіdmіnі one vіd one i souvent dans la zone de transition du corps basal vers la plaque transversale amorphe vіyu posterіgayut, à savoir yakoї pochinayutsya dans la région de la croissance axonémique des microtubules centraux. Ne vous précipitez pas. Les puants meurent et se battent. Dans tsomu rusі dіneїn є moto-and-bo avec l'écureuil rukhovy. Lors de l'association d'une dyne à des sous-unités de tubuline, on observe plus tard la formation de doublets l'un ou l'autre. Les têtes des microtubules se déplacent de l'extrémité positive à l'extrémité négative et le doublet vasculaire remonte jusqu'au sommet de la veine. Les doublets de microtubules sont liés un à un avec des protéines supplémentaires à la paire centrale de microtubules. Un tel remplacement coopératif de doublets au sommet du bik pour produire non pas au sommet de la vії, mais à la perruque. Le processus est énergivore.
Beaucoup de bactéries s'accumulent pour ruiner l'aide d'autres organes. Tsé flagelle bactérien ou flagelle. Les flagellés de bactéries sont fondamentalement viables. La puanteur peut plier Budov. Ils sont composés de trois parties principales : le long fil fibreux externe du flagelle, le corps basal et le corps basal. Le fil jgutikov a été filé à partir du flagelin blanc. Le poids moléculaire du yoga était de 40 à 60 ans. les sous-unités globulaires de la flagelline sont polymérisées en fils torsadés en spirale afin que la structure soit établie. Diamètre 12 - 25 nm. Vide au milieu. Les écureuils de la flagelline ne sont pas à la hauteur. La puanteur peut se polymériser spontanément au niveau du fil spiralé avec le chant croque de la spirale.
Près de la surface clitineuse des bactéries du flagelle, allez à la large parcelle, comme on appelle le crochet.
Dovzhina tack est proche de 45 nm. Les vins sont additionnés d'autres protéines.
Le corps basal bactérien est composé d'une cisaille attachée par une manille et d'une paire de « disques ». Un anneau est zanuren dans la membrane liposaccharidique, dans l'autre - dans la sphère de murein. Autre zanurenі dans le complexe protéique. Chez les eucaryotes, le flagelle s'effondre derrière le raton laveur du ruhu tardif des doublets. Chez les bactéries, le flagelle rux s'étend au-delà du rachnx du corps basal autour de l'axe dans le plan de la membrane plasmique. Ruh dzhgutikiv réside dans l'ATP.
La troisième partie de l'entrepôt - Filament intermédiaire 10 nm. La puanteur proviendra des monomères fibrillaires. La conception principale des filaments intermédiaires est de deviner la corde, qui peut fixer le torse.
La localisation des filaments intermédiaires est suvoro centrée sur la clitine. La puanteur se propage dans la zone nucléaire en faisceaux de fibrilles, qui remontent à la périphérie des cellules.
Les filaments intermédiaires sont piégés dans tous les types de clitines, et particulièrement riches en clitines calmes, yakі skhilnі à vplivіv mécanique. Par exemple, épiderme, m'yazi, fibres nerveuses. Il n'y a pas de filaments intermédiaires dans les clitines de roslin.
Avant l'entrepôt de filaments, un grand groupe de protéines isomères entre, qui sont subdivisées en groupes chotiri:
1) Fibres de kératine. La puanteur du bâtiment avant polymérisation. Ils sont constitués de deux sous-types. Podіlyayutsya sur aigre et neutre.
2) Vimentine, fibres de vimentine, typiques des tissus mésenchymateux. Desmine. Caractéristique pour le tissu m'yazovoi, d'ailleurs, et basané lisse et transversal. La protéine gliale est la coquille de plusieurs neurones.
3) Neurofilaments. Axones des cellules nerveuses.
4) Protéines laminaires. La puanteur ne se trouve pas dans la boule sous-membranaire du clitine, mais le reste des données a montré qu'il y a une lame et des filaments intermédiaires derrière la puissance quotidienne.
Pour toutes les protéines industrielles, une séquence d'acides aminés similaire est caractéristique, représentée par 130 acides aminés en excès dans la partie centrale de la fibrille, qui peut avoir un bourgeon en spirale - hélice alpha (la même pour toutes).
Les kіltsevі dilyanki sont caractérisés par une surface d'acides aminés différente, une dovzhina différente, non représentée par une spirale.
La présence des domaines centraux permet la création d'une sous-hélice - un dimère d'une longueur d'environ 48 nm. Les dimères associent plich-o-plich. Ils forment un court protorfilament, qui aura 4 molécules primaires et s'appelle un vin tétramère. L'épaisseur est voisine de 3 nm. Les protofilaments sont à nouveau introduits par paires et les anciennes fibrilles fines sont formées à partir de huit protofilaments ultérieurs (octamère, diamètre 10 nm). Qui possède toutes les particularités de la polymérisation des filaments intermédiaires.
Protéines de la lamina nucléaire, pue polymériser de toute façon. Les stinks font des dimères avec des têtes à un point, les stinks, polis par 2, forment un duvet avec un réseau rectangulaire. Ce type de croissance est associé à des dimères, s'accumulant jusqu'à la réaction de phosphorylation, qui conduit à la désintégration de réseaux rectangulaires dodus. Les filaments intermédiaires cytoplasmiques sont considérés comme les éléments les plus stables et les plus durables du cytosquelette. Tse i є istno-système de support.
Tsikavo, que l'expansion des filaments intermédiaires duplique l'expansion des microtubules. Lorsque le microtubule s'effondre, on voit une cigale, comme on l'appelle effondrement des filaments intermédiaires. Les odeurs sont captées par les faisceaux de fentes autour du noyau.
Fonctions des filaments intermédiaires :
1) Structurel, contre les forces de traction ;
2) Intégration de trois systèmes de clitine : l'appareil superficiel, le noyau cytosol.
Podsumok ceux-là. Dans l'entrepôt du cytosquelette, nous pouvons voir de tels éléments du cytosquelette: uniquement squelettique (filaments proximaux) et support-rukhovi (actine-myosine, tubuline-dynéine). Dans les éléments porteurs, il existe 2 manières différentes de se déplacer :
1) Établissement de la croissance de la protéine principale des microfilaments dans l'actine et de la protéine principale des microtubules de tubuline avant la polymérisation et la dépolymérisation, ce qui peut conduire à la liaison de ces protéines avec la membrane plasmique à des changements d'apparence morphologiques et à l'établissement de pseudopodes , avec la méthode de déplacement des cellules à la surface du substrat.
2) D'une autre manière, les fibrilles d'actine ou de tubuline ont été transférées par des structures directes, qui déplacent des protéines lâches spéciales - des moteurs. La puanteur interagit avec la membrane et les composants fibrillaires de la clitine, appelant au mouvement.
- La structure des structures protéiques filiformes - microtubules et microfilaments, qui forment le système de support-ruhov de la cellule.
Le cytosquelette est un système hautement dynamique du cytoplasme. De nombreuses structures du cytosquelette peuvent facilement s'effondrer et se régénérer, modifiant leur morphologie ou leur morphologie. La base de ces caractéristiques du cytosquelette est la réaction de polymérisation-dépolymérisation des principales protéines structurelles du cytosquelette et leur interaction avec d'autres protéines, à la fois structurelles et régulatrices.
Le cytosquelette ne peut être que des cellules eucaryotes, dans les cellules des procaryotes (bactéries) il n'y en a pas, ce qui est une distinction importante entre ces deux types de cellules. Le cytosquelette du clitin sing est formé pour l'apparition de la paroi dure du clitin. Vient organiser tous les organoïdes dans le cytoplasme (le soi-disant dépassement protoplasmique), qui sous-tendent la ruée amiboïde. Le cytosquelette est facile à changer, sans qu'il soit nécessaire de modifier la forme de la clitine. Le développement de la clitine modifie la forme du mouvement des couches de clitine aux premiers stades du développement embryonnaire. En cas de subdivision des cellules (mitose), le cytosquelette "se désassemble" (dissociation), et dans les cellules filles, son auto-pliage est renouvelé.
Les fonctions du cytosquelette sont différentes. Vіn spryaє podtrimtsі forme kіtini, zdіysnyuє tous les types kіtinnyh ruhіv. De plus, le cytosquelette peut participer à la régulation de l'activité métabolique des cellules.
Le cytosquelette de l'indulgence protéique. Dans le cytosquelette, on peut voir un saupoudrage des principaux systèmes, se rangeant soit derrière les principaux éléments structuraux, retenus lors des études en microscopie électronique (Microfilaments, filaments intermédiaires, microtubules), soit derrière les principales protéines, qui entrent jusqu'à leur entrepôt ( système actine-myosine, kératine, tubuline-dynéine ) ).
Filaments intermédiairesє structure la moins raisonnable parmi les principaux composants du cytosquelette pour leur repliement, leur dynamique et leurs fonctions. Leur puissance et leur dynamique sont fortement influencées par les caractéristiques des microtubules et des filaments d'actine. Les fonctions des filaments intermédiaires du dos restent dans le domaine des hypothèses.
Des filaments intermédiaires cytoplasmiques ont été révélés dans le nombre le plus important de clitines ukaryotes, à la fois chez les créatures vertébrales et sans épines, et chez les plus grands roslins. Appliquez de manière rigide les créatures de clitine, si elles ne présentent pas de filaments intermédiaires, qu'il est impossible de laisser derrière, des éclats de protéines de filaments intermédiaires peuvent former des structures non primaires.
Microtubules morphologiquesє cylindres vides d'un diamètre d'environ 25 nm avec une épaisseur de paroi d'environ 5 nm. La paroi du cylindre est composée de protofilaments - polymères linéaires de tubuline avec des hétérodimères à orientation tardive. À l'entrepôt de microtubules de protofilament, allez uzdovzh х dovgoї osі s petits dommages un à un, donc la sous-unité de tubuline utvoryut hélice à trois départs. 13 protofilaments sont inclus dans l'entrepôt de microtubules de la plupart des créatures
Filaments d'actine jouent un rôle clé dans l'appareil à mouvement rapide des m'yazovyh et des clitines non linguistiques, et participent également à d'autres processus riches en clitine, tels que la friabilité, le maintien de la forme de la clitine, la cytokinèse
Les filaments d'actine ou actine fibrillaire (actine F) sont de fines fibrilles d'un diamètre de 6 à 8 nm. La puanteur est le résultat de la polymérisation de l'actine globulaire - G-actine. Dans les cellules, les filaments d'actine à l'aide d'autres protéines peuvent former différentes structures impersonnelles.